Telmatosaurus
-
 custodian
custodian
- Вымершие животные
- 13.12.2017 00:16
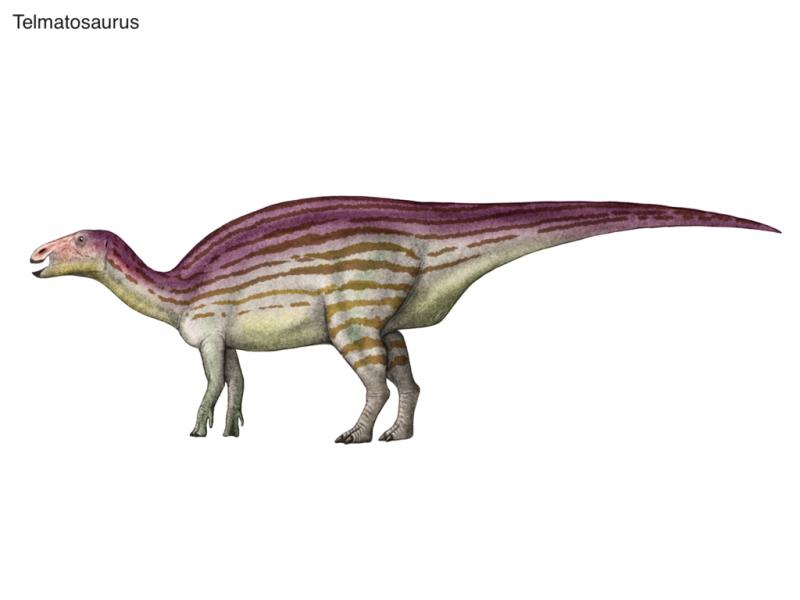
Telmatosaurus (лат. , от др.-греч. τελμᾰτιαῖος σαῦρος «болотная ящерица») — архаичный род птицетазовых динозавров, принадлежащий к семейству гадрозавридов. Считается базальным родом этого семейства. Обнаружен в отложениях маастрихтского яруса позднего мелового периода на территории современной Румынии в конце XIX века, описан австро-венгерским учёным Францем Нопчей в 1899 году. Известен по одному виду, Telmatosaurus transsylvanicus.
Весной 1895 года 12-летняя Илона Нопча, дочь местного помещика, гуляя по окрестностям деревни Сечел (ныне жудец Хунедоара, Румыния), нашла окаменевшие кости — первые в истории, обнаруженные в Трансильвании. Девочка отнесла несколько костей старшему брату, 18-летнему Францу Нопче, тем самым определив его будущую судьбу: Франц Нопча стал известным палеонтологом, «охотником за динозаврами». Найденные Илоной костные останки (обломки черепа, нижняя челюсть и несколько позвонков) стали голотипом нового рода, описанного Нопчей в 1899 году как Limnosaurus. Вскоре, однако, выяснилось, что родовое имя Limnosaurus уже занято — его в 1871 году присвоил ископаемому крокодилу Г. Ч. Марш (через сто лет родовое имя Limnosaurus было признано сомнительным, и описанные Маршем окаменелости были переклассифицированы как останки представителя рода Pristichamps). В 1903 году Нопча дал своему динозавру новое родовое имя Telmatosaurus.

От других гадрозаврид тельматозавра отличают форма зубов (не заостряющихся на конце и не имеющих характерного «гребня» вдоль внешней стороны), зазубренная предчелюстная кость и отсутствие мощного, угловатого дельтопекторального узла для крепления мышц на плечевой кости. На базальность этого рода по отношению к большим подсемействам гадрозавридов — гадрозавринам и ламбеозавринам — указывает то, что, согласно кладистическим анализам 1990-х годов он имел сходные черты с каждым из этих подсемейств. Это обстоятельство стало поводом для разногласий в отношении классификации рода: если одни исследователи включают его, так же как роды Tethyshadros и Bactrosaurus, в семейство гадрозавридов, но оставляют за пределами подсемейств гадрозаврин и ламбеозаврин, которых в свою очередь объединяют в общую кладу эугадрозавров (лат. Euhuadrosauria). Другие же выносят эти роды за пределы семейства гадрозавридов, оставляя в более широком таксоне гадрозавроидов, в который входят все динозавры, более близкие к гадрозавридам, чем к игуанодонтидам. Внешнее по отношению к гадрозавринам и ламбеозавринам положение данного года на эволюционном дереве постулировалось с 1950-х годов, когда Фридрих фон Хюне включал его, вместе с ещё одним базальным родом Tanius, в отдельное от гадрозавридов семейство надсемейства Hadrosauria. В то же время, с 1970-х годов до настоящего времени существует и точка зрения, согласно которой Telmatosaurus относится к ламбеозавринам.

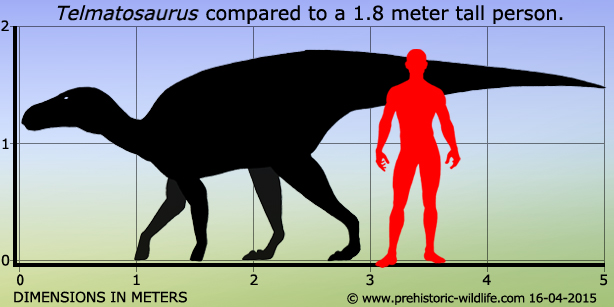
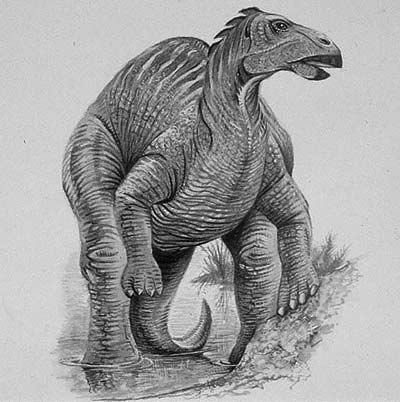
Telmatosaurus был совсем небольшим для гадрозаврида динозавром — длина его тела вместе с хвостом составляла около 5 метров, а масса тела около 500 кг, что в десять раз меньше средней массы гадрозаврид этой эпохи. Существует мнение, что эти малые размеры представляют собой пример островной карликовости, поскольку местом обитания представителей этого рода был остров Хацег — один из островов тогдашнего Европейского архипелага (другим «карликом» этого региона был также открытый Нопчей титанозаврид Magyarosaurus). Передвигался предположительно на всех четырёх ногах, при беге поднимаясь на задние ноги и развивая максимальную скорость в 15—20 км/ч, но обычно двигаясь значительно медленнее. Вероятно, использовал передние конечности, чтобы подтягивать к пасти листья и побеги, которые, предположительно, поедал на высоте не более 1 метра от поверхности.
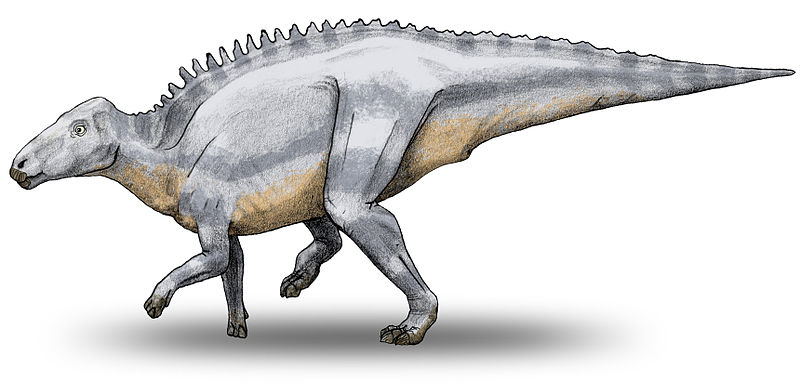
Череп тельматозавра был вытянутым, «лошадиным» и в целом напоминал как череп игуанодонов, так и черепа других гадрозавридов. Предчелюстная кость была узкой и беззубой и, по всей видимости, поддерживала кератиновый клюв. Зубной аппарат напоминает зубы других гадрозавридов — зубные «батареи» представляют собой комплекс из колонн постоянно сменяющихся на протяжении жизни зубов, способных перетирать самую жёсткую растительную пищу. Как верхняя, так и нижняя челюсть были оснащены более чем 30 зубными колоннами, что больше, чем у игуанодонов, но меньше, чем у других гадрозавридов. Хотя у тельматозавра не было специфичного для ламбеозавринов сплошного или полого костяного гребня на голове, реконструкция черепа указывает на наличие изогнутых костных наростов, идущих по сторонам морды. Эти наросты заметно отличают голову тельматозавра от голов более примитивных орнитоподов. Вейсхампел и Жиану в книге «Трансильванские динозавры» предполагают, что эти наросты, как и у других гадрозавридов, могли выполнять социальную функцию, но не могут утверждать, что это была форма полового диморфизма.
Как и другие гадрозавриды, Telmatosaurus обладал длинным и жёстким, укреплённым оссифицированными сухожилиями хвостом, который служил балансиром при движении. Хотя по сравнению с другими гадрозавридами и игуанодонтидами, чья длина зачастую превышала 10 метров, Telmatosaurus был маленьким и изящным, опора преимущественно на задние конечности при почти горизонтальном положении тела делала наличие такого балансира необходимым.
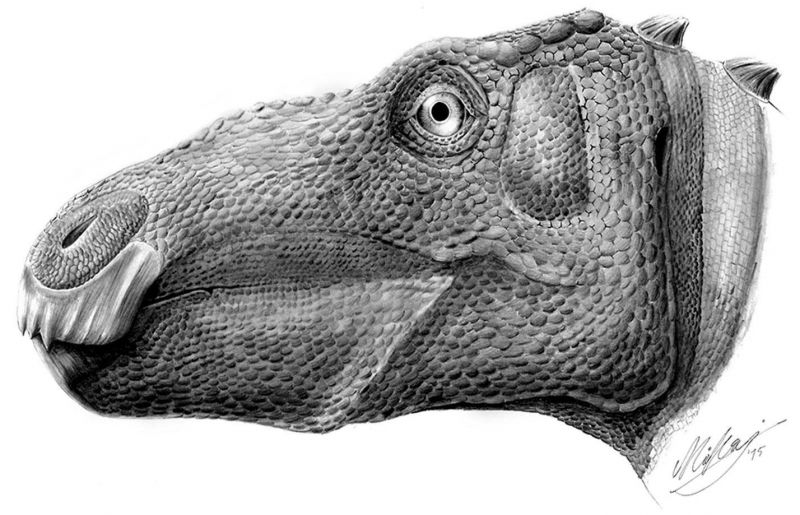
Молодой Telmatosaurus, исследованный Dumbrava и другими, имеет большую не раковую опухоль, называемую амелобластомой, на нижней челюсти. Наличие этой доброкачественной опухоли у динозавра является первым, так как до этого открытия амелобластомы были известны только у современных млекопитающих (включая людей) и у рептилий. Известно, что Telmatosaurus является базальным гадрозавридом, и поэтому наличие такой деформации в этой точке дает нам дополнительные доказательства того, что утконосые динозавры, более подвержены опухолям, чем другие. На рисунке ниже представлена реконструкция Telmatosaurus с деформацией челюсти.
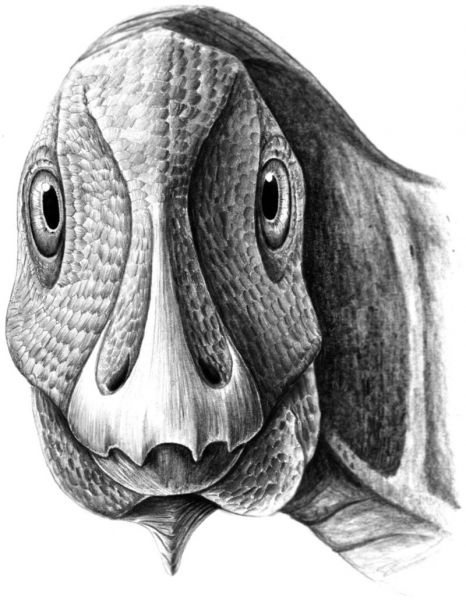
Маловероятно, что опухоль вызвала у динозавра серьезную боль на ранних стадиях развития, как и у людей с такимм же состоянием, но исследователи могут сказать по его размерам, что этот динозавр умер до того, как он достиг взрослого возраста. Поскольку сохранившиеся останки состоят только из двух нижних челюстей, нельзя определить причину смерти. Тем не менее исследователям было интересно узнать, способствовало ли присутствие амелобластомы его смерти. Из современных примеров мы знаем, что хищники часто нападают на члена стада, который выглядит немного иначе или даже слегка ослабленным от болезни. Опухоль у этого динозавра не развилась в полной мере в момент его смерти, но это могло косвенно способствовать его ранней кончине.
Источники:
https://liliensternus.deviantart.com/art/Telmatosaurus-transylvannicus-152792566
https://cisiopurple.deviantart.com/art/Telmatosaurus-577554255
https://hyrotrioskjan.deviantart.com/art/Telmatosaurus-215293915
https://en.wikipedia.org/wiki/Telmatosaurus
http://www.prehistoric-wildlife.com/species/t/telmatosaurus.html
https://ru.wikipedia.org/wiki/Telmatosaurus
http://dinodata.de/animals/dinosaurs/pages_t/telmatosaurus.php
http://www.nhm.ac.uk/discover/dino-directory/telmatosaurus.html#
Похожие статьи
-
 Eolambia
Eolambia (лат.) — род растительноядных динозавров из клады Neoiguanodontia группы игуанодонтов, живших в меловом периоде (105,3—93,5 млн лет назад) на территории современных США.
Eolambia
Eolambia (лат.) — род растительноядных динозавров из клады Neoiguanodontia группы игуанодонтов, живших в меловом периоде (105,3—93,5 млн лет назад) на территории современных США. -
 Tanius
Таний (лат. Tanius) — род травоядных орнитоподовых динозавров надсемейства Hadrosauroidea из мела Азии. Типовой вид Tanius sinensis назван и описан Wiman в 1929 году. Таний —...
Tanius
Таний (лат. Tanius) — род травоядных орнитоподовых динозавров надсемейства Hadrosauroidea из мела Азии. Типовой вид Tanius sinensis назван и описан Wiman в 1929 году. Таний —... -
 Tethyshadros
Tethyshadros (лат.) — род травоядных орнитоподовых динозавров надсемейства Hadrosauroidea из верхнемеловой эпохи Европы. Типовой и единственный вид Tethyshadros insularis...
Tethyshadros
Tethyshadros (лат.) — род травоядных орнитоподовых динозавров надсемейства Hadrosauroidea из верхнемеловой эпохи Европы. Типовой и единственный вид Tethyshadros insularis... -
 Protohadros
Protohadros (лат.) — род травоядных орнитоподовых динозавров надсемейства Hadrosauroidea из мела Северной Америки. Типовой и единственный вид Protohadros byrdi назван и описан ...
Protohadros
Protohadros (лат.) — род травоядных орнитоподовых динозавров надсемейства Hadrosauroidea из мела Северной Америки. Типовой и единственный вид Protohadros byrdi назван и описан ...







