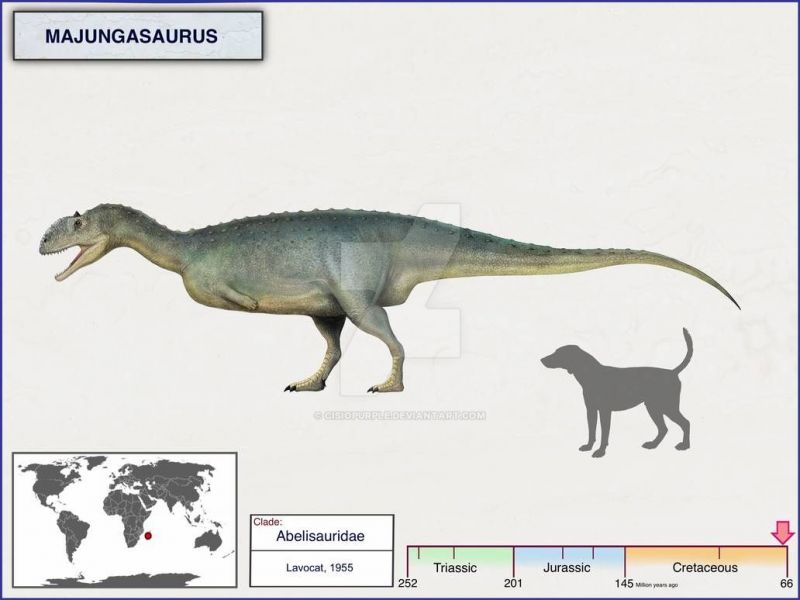
Majungasaurus
-
 custodian
custodian
- Вымершие животные
- 13.04.2020 20:17

Майюнгазавр (лат. Majungasaurus, «ящер из Махадзанги») — род тероподовых динозавров из семейства абелизаврид, чьи ископаемые остатки найдены в слоях маастрихтского яруса верхнемеловой системы (72,1—66,0 млн лет назад) на территории Мадагаскара. В род включают типовой и единственный вид Majungasaurus crenatissimus.
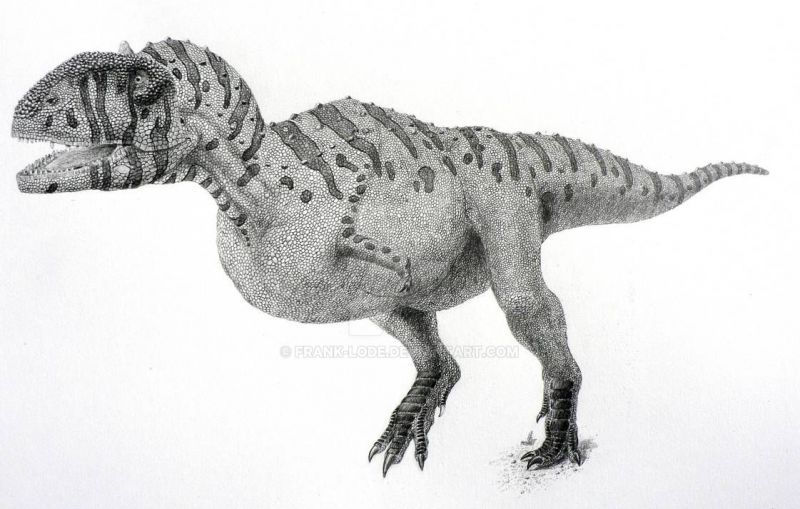
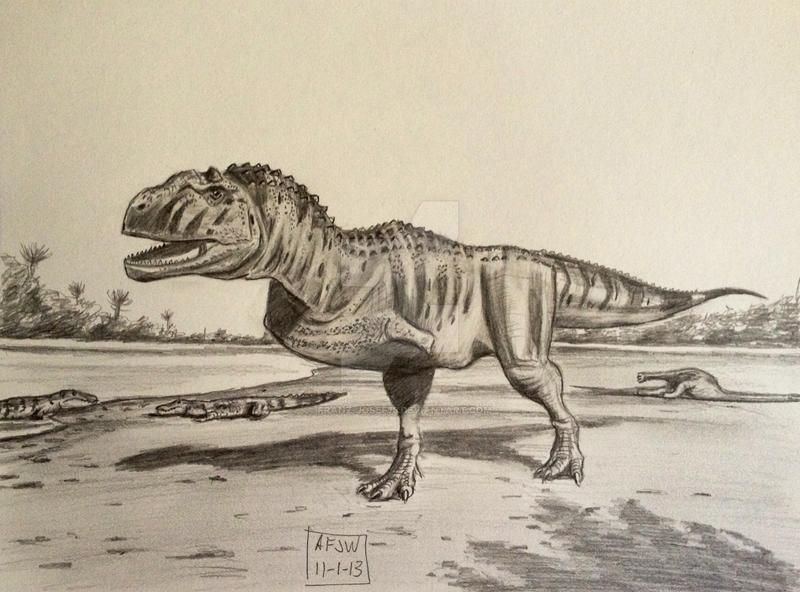
Как и прочие абелизавриды, майюнгазавр был среднеразмерным бипедальным хищником с короткой мордой. Хотя его передние конечности полностью не известны, они были очень маленькими, а задние были короткими и коренастыми. Этого динозавра можно отличить от других абелизаврид по более широкому черепу, очень грубой текстуре утолщённой кости на вершине морды, а также по одному скруглённому рогу на крышке черепа, который изначально был принят за купол пахицефалозавра. У него также было больше зубов на верхней и нижней челюстях, чем у большинства представителей семейства.
Известный по нескольким хорошо сохранившимся черепам и множественному скелетному материалу, майюнгазавр стал одним из наиболее изученных теропод Южного полушария. Похоже, что он более тесно связан с абелизавридами из Индии, чем из Южной Америки или континентальной Африки, и этот факт имеет важные биогеографические последствия. Майюнгазавр был сверххищником в своей экосистеме, который охотился в основном на завропод, таких как рапетозавры, а также является одним из немногих динозавров, для которых есть прямые доказательства каннибализма.

Найденные в 1896 году французским палеонтологом Шарлем Депере два зуба и несколько позвонков считаются первыми окаменелостями хищных динозавров на Мадагаскаре. Депере отнес эти остатки к новому виду Megalosaurus crenatissimus. Многочисленные фрагментарные остатки находили в течение следующих 100 лет. В 1955 году французский палеонтолог Рене Лавокат на основании новых зубов описал новый род Majungasaurus — «ящер из Маюнги». Позже в 1979 году в тех же отложениях были обнаружены куски утолщенных лобных костей какого-то динозавра. Находку описал Филипп Тагует как Majungatholus atopus. Считалось, что это остатки большого пахицефалозавра.

В 1996 году был обнаружен череп, сочетавший зубы маюнгазавра и утолщенные лобные кости маюнгатолуса, который после изучения был отнесен к абелизавридив. Далее в 2003-2005 годах было найдено множество костей по которым можно судить о строении скелета этого динозавра. С 2007 года все окаменелости принадлежат к роду Majungasaurus crenatissimus.
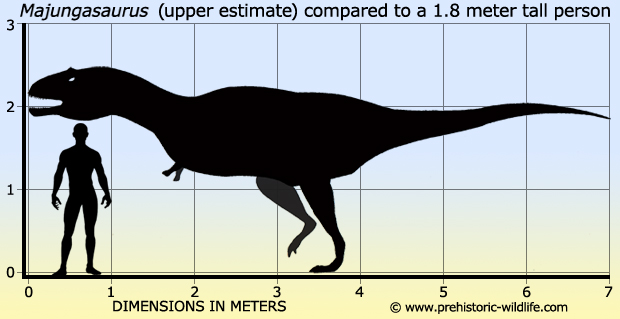
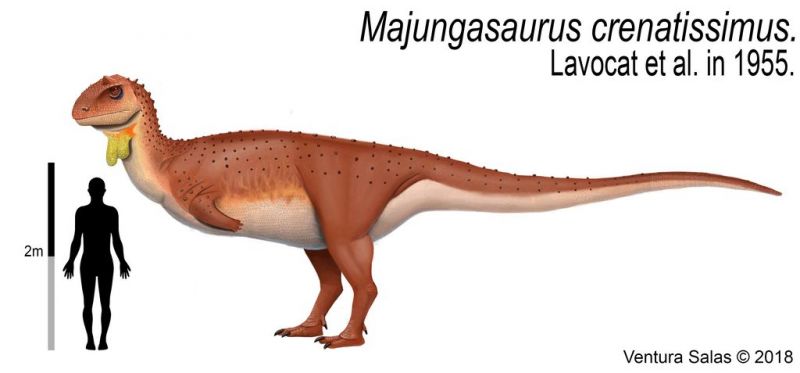
Майюнгазавр был тероподом среднего размера, длина которого составляла 6—7 метров. Фрагментарные остатки крупных особей указывают, что некоторые взрослые животные могли достигать длины 8—11 метров. Грегори С. Пол оценил массу динозавра в 750 килограммов, Сэмпсон и Уитмер дали более внушительную оценку — 1100 килограммов. Однако, образец FMNH PR 2100, на котором они провели измерения, был не самым большим из найденных. Более крупные экземпляры майюнгазавра сравнимы по размеру с родственным карнотавром, который, по оценкам, весил 1500 килограммов.

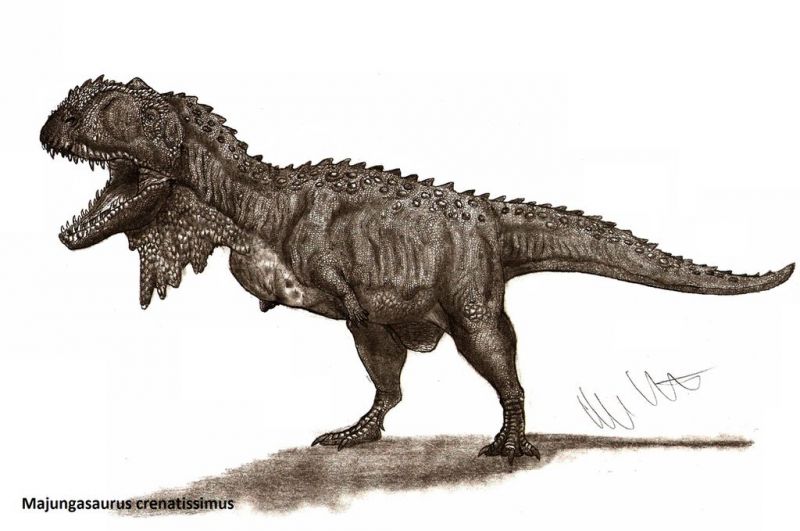
Череп майюнгазавра известен исключительно хорошо по сравнению с большинством теропод и в целом похож на черепа других представителей семейства. Как и у других абелизаврид, его длина пропорционально короткая относительно высоты, хотя и не такая короткая, как у карнотавра. Черепа крупных особей имели длину 60—70 сантиметров. Типичной для семейства является высокая предчелюстная кость, которая визуально сильно притупляла кончик морды. Тем не менее, череп майюнгазавра был заметно шире черепов других абелизаврид. Черепа всех абелизаврид имеют грубую скульптурированную поверхность наружных граней костей, и майюнгазавр не является исключением. Эта анатомическая особенность доведена у него до крайности: его носовые кости были чрезвычайно толстыми, сросшимися с низким центральным гребнем, проходящим вдоль ноздрей. Отличительный куполообразный рог выступал из слитых лобных костей на верхней части черепа. В жизни эти структуры были покрыты неким покровом, возможно, состоящим из кератина. Компьютерная томография черепа показала, что как рог, так и носовая структура содержат пазухи, возможно, служившие для уменьшения веса головы.

Посткраниальный скелет майюнгазавра очень схож со скелетами карнотавра и Aucasaurus — единственных абелизаврид с полным известным скелетным материалом. Майюнгазавр был бипедальным животным с длинным хвостом, уравновешивающим голову и туловище, с центром масс в районе бёдер. Хотя шейные позвонки содержали многочисленные полости (плевроцели) для уменьшения веса, они были крепкими, с увеличенными участками крепления мышц, и шейными рёбрами для силы. К шейным рёбрам крепились окостенелые сухожилия, придавая им раздвоенный вид — точно так же, как у карнотавра. Все эти особенности делали шею животного очень сильной и мускулистой. Уникальной особенностью шейных рёбер майюнгазавра были длинные углубления по сторонам для снижения веса. Плечевая кость была короткой и изогнутой, очень похожей на таковую у карнотавра и Aucasaurus. Так же, как и у родственных видов, у майюнгазавра были очень короткие передние конечности с четырьмя сильно редуцированными пальцами, состоящими из одной (I и IV пальцы) или двух (II и III пальцы) фаланг. На костях плюсны и пальцев у майюнгазавра, как и прочих представителей подсемейства Majungasaurinae, отсутствовали характерные ямки и бороздки, к которым крепились когти и сухожилия, а кости пальцев были слиты воедино, что указывает на то, что кисть животного была неподвижной. В 2012 году был описан образец лучшей сохранности, показывающий, что нижняя часть передней конечности была крепкой, хотя и короткой, и что кисть состояла из 4 плюсневых костей и 4, вероятно, несгибающихся и сильно редуцированных пальцев с небольшими когтями на II и III пальцах.

Как и у других абелизаврид, задние конечности майюнгазавра были коренастыми и пропорционально короткими по сравнению с длиной туловища. Его большеберцовая кость была ещё более короткой, чем у карнотавра, с заметным гребнем на колене. Таранная и пяточная кости были слиты воедино, а на ногах было 3 функциональных пальца и маленький I палец, который не касался земли.

Пожалуй, самой характерной чертой майюнгазавра была орнаментация черепа: выпуклые, слитые носовые кости и рог на лбу. Другие представители группы цератозавров, такие как карнотавр, цератозавр (лат. Ceratosaurus) и Rajasaurus, несли на голове гребни. Эти костные структуры, вероятно, играли роль во внутривидовой конкуренции, хотя их точная функция в этом контексте неизвестна. Полость лобного рога майюнгазавра ослабляла эту структуру и, вероятно, исключала её применение в прямом физическом бою, хотя рог, возможно, служил в демонстрационных целях. В то время как есть различия в орнаментации разных особей майюнгазавра, нет никаких доказательств полового диморфизма у этих животных.
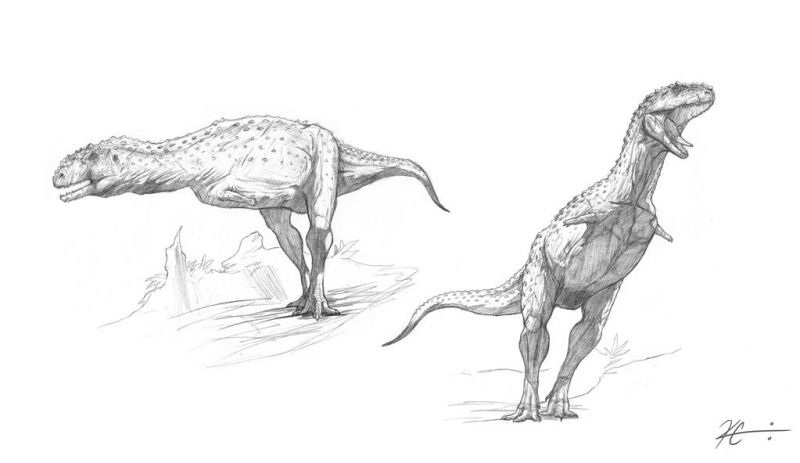
Учёные предположили, что уникальная форма черепа майюнгазавра и других абелизаврид указывает на несколько иные хищнические привычки, чем у прочих теропод. В то время как большинство теропод характеризовалось длинными, низкими и узким черепами, черепа абелизаврид были выше, шире и короче. Узкие черепа других теропод имели строение, позволяющее противостоять вертикальному напряжению, возникающему при укусе, но не так хорошо выдерживали скручивание. Если сравнивать с современными хищными млекопитающими, большинство теропод, вероятно, использовало стратегию, схожую в некоторой степени с таковой у длинно- и узкомордых псовых, которые многочисленными укусами ослабляют свою жертву.

Вместо этого абелизавриды, и особенно майюнгазавр, могли использовать стратегию охоты, сходной с таковой у современных кошачьих с короткими и широкими мордами, которые кусают один раз и удерживают добычу, пока она не ослабеет. У майюнгазавра ещё более широкая морда, чем у прочих абелизаврид, и в пользу гипотезы укуса и удержания могут говорить другие аспекты его анатомии. Его шея была усилена крепкими позвонками, переплетёнными шейными рёбрами и окостеневшими сухожилиями; помимо этого, места прикрепления мышц к шейным позвонкам и к задней стороне черепа также были очень сильными. Черепа абелизаврид также были усилены минерализованными костями, выступавшими за поверхность кожи и создававшими характерную грубую текстуру головы. Особенно хорошо это видно у майюнгазавра, чьи носовые кости слиты и утолщены. С другой стороны, нижняя челюсть этого динозавра имела большие окна с каждой стороны, как и у других цератозавров, а также синовиальные суставы между определёнными костями, которые обеспечивали высокую степень гибкости нижней челюсти, хотя и не такую высокую, как у змей. Такие особенности анатомии могли служить для предотвращения перелома челюсти при удержании сопротивляющейся жертвы. Передние зубы верхней челюсти были более крепкими, чем остальные, и обеспечивали опорную точку для укуса, в то время как низкие коронки майюнгазавра препятствовали ломке зубов во время борьбы. Наконец, в отличие от зубов аллозавра и большинства других теропод, которые были изогнуты как спереди, так и сзади, у абелизаврид были зубы, изогнутые на передней кромке, но более прямые на задней (режущей) кромке. Благодаря такому строению зубы хищника крепко вцеплялись в плоть жертвы, не разрывая мясо на куски. Исследование зубов майюнгазавра показывает, что его зубы менялись от 2 до 13 раз чаще, чем у других теропод, заменяя весь зубной ряд в течение 2 месяцев. Причиной для такой быстрой смены зубов, возможно, служила привычка животного разгрызать кости.

В своей экосистеме майюнгазавры были сверххищниками, а единственными крупными растительноядными животными были завроподы, такие как рапетозавры. Учёные предположили, что майюнгазавры, и, возможно, прочие абелизавриды специализировались на охоте на завропод. Анатомические адаптации головы и шеи для стратегии «укус и удержание» могли быть полезны против завропод, которые были очень сильными животными. В пользу этой гипотезы говорят короткие и коренастые задние конечности хищника, отличные от более длинных и тонких ног большинства других теропод. Хотя такие ноги не позволяли майюнгазавру двигаться так же быстро, как другие тероподы, это не создавало для него проблем в преследовании медлительных завропод. Прочные кости задних конечностей предполагают очень сильные ноги, а их более короткая длина могла бы понизить центр тяжести животного. Таким образом, майюнгазавр, возможно, жертвовал скоростью ради силы. Следы зубов майюнгазавра на костях рапетозавра подтверждают, что он, по крайней мере, питался этими завроподами независимо от того, действительно ли он их убивал.
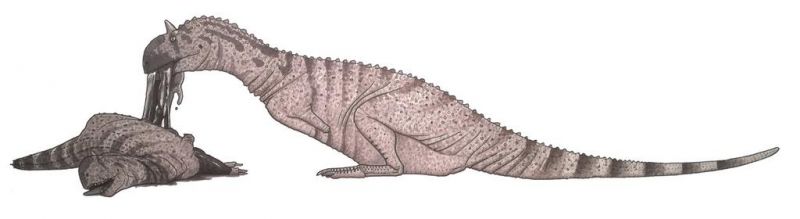
Несмотря на то, что завроподы были основой рациона майюнгазавра, существуют находки, говорящие в пользу присутствия в их диете особей своего вида. Было обнаружено множество костей майюнгазавра с отметинами зубов, аналогичными найденным на костях завропод в тех же местах. Эти отметины имеют тот же размер, то же расстояние и несут более мелкие царапины, соответствующие пильчатой насечке на зубах майюнгазавра. Поскольку майюнгазавры — единственные крупные тероподы Мадагаскара, самое простое объяснение состоит в том, что они питались другими особями своего вида. Предположения о том, что триасовый теропод целофиз (лат. Coelophysis) был каннибалом, опровергнуты, в результате чего майюнгазавр стал единственным динозавром с подтверждёнными склонностями к поеданию себе подобных, хотя имеются некоторые свидетельство существования каннибализма у других видов.

Неизвестно, активно ли охотился майюнгазавр на особей своего вида или только обгладывал из туши. Однако некоторые исследователи отмечают, что современные комодские вараны иногда убивают друг друга, когда соревнуются за доступ к мёртвым телам жертв. После этого вараны продолжают поедать остатки своих соперников, что может указывать на подобное поведение у майюнгазавра и других теропод.

Исследование, которое провели Мишель Д'Эмик и его коллеги, показало, что майюнгазавр был одним из самых медленно растущих теропод. Изучив линии роста нескольких костей, учёные установили, что этому динозавру требовалось около 20 лет — большая часть жизни — для достижения максимального размера. Животное, чей образец изучили палеонтологи, к 27 годам достигло длины в 6 метров и массы 850 килограммов. Для примера, тираннозавр набирал 800 килограммов всего за несколько лет, достигая массы в 8—9 тонн к концу жизни. Такой медленный рост животного, вероятно, был обусловлен суровыми условиями жизни: климат мелового Мадагаскара изобиловал засухами и наводнениями. Однако было обнаружено, что и другие абелизавриды также имеют сравнительно медленные темпы роста, что могло быть общей особенностью группы.

Все образцы майюнгазавра были извлечены из геологической формации Мейварано на северо-западе Мадагаскара. Большая их часть, включая наиболее сохранившиеся скелеты, были найдены в стратиграфическом горизонте Анембалемба, хотя зубы этого динозавра были также обнаружены в подстилающем горизонте Масоробе и перекрывающем горизонте Миадана. Несмотря на то, что эти отложения не подвергались радиометрической датировке, данные биостратиграфии и палеомагнетизма предполагают, что они образовались во время маастрихтской эпохи, которая длилась от 72 до 66 миллионов лет. Зубы майюнгазавра находят до самого конца маастрихта, вплоть до мел-палеогенового вымирания.
В позднемеловую эпоху, как и сейчас, Мадагаскар был островом, отколовшимся от Индийского субконтинента за 20 миллионов лет до этого. Он дрейфовал на север, находясь всё ещё на 10—15 ° южнее, чем сегодня. Преобладающий климат того времени был полузасушливым, с выраженной сезонностью по температуре и количеству осадков. Майюнгазавры населяли прибрежную пойму, прорезанную множеством песчаных речных каналов. Убедительные геологические данные говорят о том, что в начале сезона дождей эти каналы периодически заполнялись потоками мусора, закапывая туши организмов, погибших в предшествующий сухой сезон и обеспечивая их исключительную сохранность в виде окаменелостей. Уровень моря в этом районе повышался на протяжении всего маастрихта и будет продолжаться в эпоху палеоцена, поэтому майюнгазавр, возможно, также бродил по прибрежным районам, таким, как приливные равнины. Соседняя формация Беривотра представляет существовавшую в то же время морскую среду.
Помимо майюнгазавра, ископаемые таксоны, извлечённые из отложений формации Мейварано, включают рыб, лягушек, ящериц, змей, 7 различных видов крокодиломорфов, 5 или 6 видов млекопитающих, птиц, таких, как Vorona, возможно летающего дромеозаврида Rahonavis, ноазаврида Masiakasaurus и двух титанозавровых завроподов, включая репетозавра (лат. Rapetosaurus). Майюнгазавры, безусловно, были крупнейшими сухопутными хищниками Мадагаскара, хотя некоторые крокодиломорфы, такие как Mahajangasuchus и Trematochampsa, могли конкурировать с ним ближе к воде.
На рисунке ниже два майюнгазавра охотятся на рапетозавра.

На рисунке ниже самец Majungasaurus пытается привлечь самку.

На рисунке ниже Majungasaurus рассматривает Rahonavis присевшего ему на спину.

Источники:
http://www.prehistoric-wildlife.com/species/m/majungasaurus.html
https://ru.wikipedia.org/wiki/%D0%9C%D0%B0%D0%B9%D1%8E%D0%BD%D0%B3%D0%B0%D0%B7%D0%B0%D0%B2%D1%80
https://uk.wikipedia.org/wiki/Majungasaurus_crenatissimus
https://en.wikipedia.org/wiki/Majungasaurus
https://www.deviantart.com/spinoinwonderland/art/Majungasaurus-crenatissimus-632096245
https://www.deviantart.com/frank-lode/art/Majungasaurus-386821533
https://www.deviantart.com/green-mamba/art/006-MAJUNGASAURUS-CRENATISSIMUS-280633078
https://www.deviantart.com/darthmobius/art/Majungasaurus-717427231
https://www.deviantart.com/teratophoneus/art/Majungasaurus-Crenatissimus-515472123
https://www.deviantart.com/malvit/art/Crowned-Majungasaurus-All-Yesterdays-358356859
https://www.deviantart.com/c-compiler/art/Majungasaurus-Courtship-507441614
https://www.deviantart.com/tnilab-ekneb121/art/Majungasaurus-portrait-815223937
https://www.deviantart.com/paleocolour/art/Majungasaurus-crenatissimus-572692577
https://www.deviantart.com/kevcatalan/art/Majungasaurus-451224971
https://www.deviantart.com/c-compiler/art/Male-Majungasaurus-closeup-507449305
https://www.deviantart.com/christopher252/art/Majungasaurus-crenatissimus-687120852
https://www.deviantart.com/zegh8578/art/Majungasaurus-673704000
https://www.deviantart.com/josefavaldiviat-rex/art/Majungasaurus-crenatissimus-running-612083537
https://www.deviantart.com/ntamura/art/Majungasaurus-84968304
https://www.deviantart.com/cisiopurple/art/Majungasaurus-757825938
https://www.deviantart.com/franz-josef73/art/Malagasy-Marauder-Majungasaurus-410840937
https://www.deviantart.com/venturasalas/art/Majungasaurus-crenatissimus-763766693
https://dinosaurpictures.org/Majungasaurus-pictures
https://nsf.gov/news/mmg/mmg_disp.jsp?med_id=73607
https://www.nhm.ac.uk/discover/dino-directory/majungasaurus.html
https://www.deviantart.com/durbed/art/Majungasaurus-crenatissimus-280469594
https://www.deviantart.com/paleoguy/art/Majungasaurus-Mating-Display-673984703

Похожие статьи
-
 Rahiolisaurus
Рахиолизавр (лат. Rahiolisaurus, буквально — ящер из Рахиоли) — род тероподовых динозавров из семейства абелизаврид, живших в позднем меловом периоде (маастрихт) около 70—65...
Rahiolisaurus
Рахиолизавр (лат. Rahiolisaurus, буквально — ящер из Рахиоли) — род тероподовых динозавров из семейства абелизаврид, живших в позднем меловом периоде (маастрихт) около 70—65... -
 Rajasaurus
Раджазавр (лат. Rajasaurus narmadensis) — хищный динозавр конца позднемеловой эпохи (маастрихт). Принадлежит к группе абелизавров. Описан в 2003 году на основании остатков,...
Rajasaurus
Раджазавр (лат. Rajasaurus narmadensis) — хищный динозавр конца позднемеловой эпохи (маастрихт). Принадлежит к группе абелизавров. Описан в 2003 году на основании остатков,... -
 Spectrovenator
Spectrovenator — род хищных абелизавровых динозавров, существовавший в раннем меловом периоде. Почти полный скелет найден близ городка Корасан-ди-Жезус, Бразилия. Первый...
Spectrovenator
Spectrovenator — род хищных абелизавровых динозавров, существовавший в раннем меловом периоде. Почти полный скелет найден близ городка Корасан-ди-Жезус, Бразилия. Первый... -
 Quilmesaurus
Куилмезавр (лат. Quilmesaurus) — род плотоядных динозавров из верхнего мела из семейства абелизаврид, живший на территории современной Аргентины. Единственными известными...
Quilmesaurus
Куилмезавр (лат. Quilmesaurus) — род плотоядных динозавров из верхнего мела из семейства абелизаврид, живший на территории современной Аргентины. Единственными известными...







