Glanosuchus
-
 custodian
custodian
- Вымершие животные
- 30.05.2018 19:55
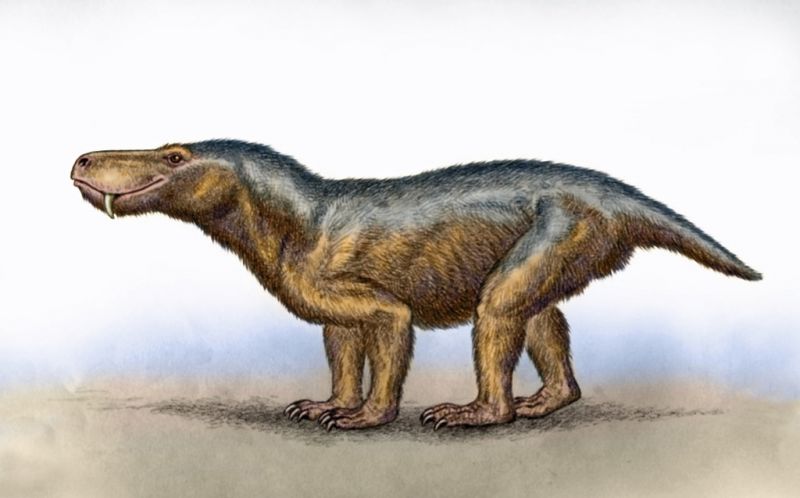
Glanosuchus — род тероцефалов из семейства Scylacosauridae, известный из поздней перми Южной Африки. Типовой вид G. macrops был назван Робертом Брумом в 1904 году. Структура среднего уха у Glanosuchus была промежуточной между ранними терапсидами и млекопитающими. Выступы в полости носа Glanosuchus предполагают, что он имел, по крайней мере, частично эндотермический метаболизм, напоминающий современных млекопитающих.
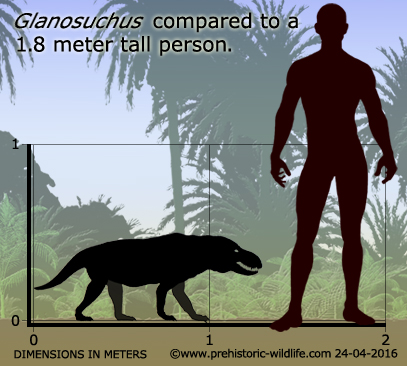
Glanosuchus macrops впервые был описан в 1904 году южноафриканским палеонтологом Робертом Брумом, который назвал род и вид на основе почти полного голотипового черепа. Череп был искажен во время окаменения, и в некоторых частях кость неотличима от окружающей матрицы. При иллюстрации голотипа Брум предпочел реконструировать череп вида, а не нарисовать фактический образец.
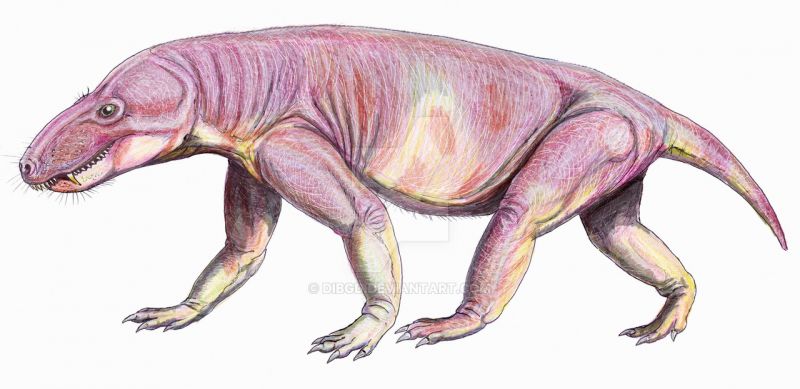
Череп Glanosuchus имеет длину около 30 см. Вероятно, Glanosuchus выростал до 1,8 м в длину. Как и у других ранних тероцефалов, у Glanosuchus была длинная, глубокая морда и большие клыки. Зубы резца на передней части верхней челюсти также большие и лопастные. Есть шесть резцов с обеих сторон верхней челюсти, самый дальний из которых заметно меньше остальных. За каждым клыком расположены пять маленьких острых зубов. Морда шире спереди, чем сзади, обычная вещь среди терапсид, но присутствующая у нескольких других родственных тероцефалов. Ноздри расположены на кончике морды и направлены вперед.
Glanosuchus представляет собой раннюю стадию развития среднего уха у млекопитающих. Современные млекопитающие имеют три кости в среднем ухе (молоточек, наковальня и стремечко), которые передают звуковую энергию от барабанной перепонки к жидкости внутреннего уха. Молоточка и наковальня млекопитающих развились из сустава и квадратной кости ранних терапсид. Изучение костей Glanosuchus показывает, что у него была очень тонкая костяная пластинка, которая действовала как барабанная перепонка, получая звуки и передавая их в небольшую заполненную воздухом полость. Стремечко и вестибулярные отверстия (отверстия, соединяющия среднее и внутреннее ухо) сохранились в одном экземпляре Glanosuchus, который был исследован путем проведения поперечных сечений черепа. Акулярная связка, кольцеобразная структура, которая образует уплотнение между концом стремечка и острием вестибулярного отверстия, вероятно, удерживалась на месте хрящами. Передача звука между тонкой костной пластиной и вестибулярным отверстием у Glanosuchus была не такой эффективной, как у млекопитающих, а это означало, что у животного было менее острое чувство слуха.
Glanosuchus, возможно, был одним из первых терапсид, которые достигли эндотермии или теплокровности. Эндотермия наблюдается сегодня у млекопитающих, единственной существующей группы терапсид. Рептилии, самые близкие живые родственники млекопитающих, являются хладнокровными эктотермами с более низким уровнем метаболизма. Эндотермические животные, вероятно, эволюционировали из более примитивных эктотермических синапсидов в пермском или триасовом.
В то время как мех, обычно принимаемый как четкое указание на эндотермию, не был обнаружен у ранних терапсид, некоторые скелетные признаки, сохранившиеся в остатках терапсид, могут быть признаком скорости метаболизма этих животных. Современные млекопитающие обладают носовыми раковинами в полости носа, которые собирают влагу из вдыхаемого воздуха. В качестве эндотерм млекопитающие должны быстро дышать, чтобы обеспечить достаточное количество кислорода для своего высокого метаболизма. По мере поступления кислорода в полость носа и из него он высушивает окружающие ткани. Вода из вдыхаемого воздуха конденсируется на этих носовых раковинах, предотвращая высыхание носовой полости и позволяя млекопитающим вдыхать достаточное количество кислорода для поддержания их высокого метаболизма.
Рептилии и более примитивные синапсиды имеют носовые раковины, но эти пластины кости участвуют в ощущении запаха, а не в предотвращении высыхания. В то время как носовые раковины млекопитающих расположены на пути потока воздуха для сбора влаги, сенсорные раковины как у млекопитающих, так и у пресмыкающихся расположена дальше назад и над носовым проходом, подальше от потока воздуха. Glanosuchus имеет выступы, расположенные низко в носовой полости, что указывает на то, что у него были нижние носовые раковины, находящиеся на прямом пути воздушного потока. Возможно, верхние носовые раковины не сохранились, потому что они были либо очень тонкими, либо хрящевыми. Также была высказана мысль о том, что эти раковины связаны с обонятельным эпителием, а не с носовыми раковинами. Тем не менее, возможное присутствие носовых раковин предполагает, что Glanosuchus, возможно, был в состоянии быстро дышатьбез пересыхания носового прохода, а, следовательно, может быть теплокровным.
Glanosuchus — это самый ранний известный терапсид, обладающий носовыми раковинами, но он разделяет некоторые черты с рептилиями, что предполагает, что он не был полностью теплокровным. Хоаны, два отверстия в небе, которые соединяют носовую полость с ртом, расположены далеко впереди у пресмыкающихся, ранних синапсид и Glanosuchus. Это уменьшает полость носа, тем самым уменьшая способность увлажнять входящий воздух. В дальнейшей эволюции тероцефалов хоаны мигрировали дальше по небу, указывая на то, что у таких продвинутых форм, как Bauria, были высокие показатели метаболизма, подобные современным млекопитающим. Когда хоаны двигались дальше назад, перед ними расширялось вторичное небо. Это расширение имело место как у тероцефалов, так и у родственных цинодонтов, что указывает на то, что две группы одновременно получали признаки млекопитающих в перми и триасе. Несмотря на то, что тероцефалы вымерли в среднем триасе, цинодонты продолжали диверсифицироваться, дав начало полностью теплокровным млекопитающим в позднем триасе.
Источники:
https://willemsvdmerwe.deviantart.com/art/Glanosuchus-macrops-746165127
https://mojcaj.deviantart.com/art/Glanosuchus-120993327
https://dibgd.deviantart.com/art/Glanosuchus-116755546
http://www.prehistoric-wildlife.com/species/g/glanosuchus.html
https://twitter.com/synapsida/status/502819348785168384
http://1cperm.ru/fauna/as-scylacos.php
https://en.wikipedia.org/wiki/Glanosuchus

Похожие статьи
-
 Пристерогнат
Пристерогнат (лат. Pristerognathus, от др.-греч. πριστήρ γνάθος «распиливающие челюсти») — примитивный тероцефал пермского периода. Происходит из среднепермских отложений...
Пристерогнат
Пристерогнат (лат. Pristerognathus, от др.-греч. πριστήρ γνάθος «распиливающие челюсти») — примитивный тероцефал пермского периода. Происходит из среднепермских отложений... -
 Поростеогнат
Поростеогнат (Porosteognathus efremovi) — среднепермский тероцефал из Ишеевской фауны нижнетатарского подъяруса в Татарстане. Известен по фрагментам черепов, описан Б. П....
Поростеогнат
Поростеогнат (Porosteognathus efremovi) — среднепермский тероцефал из Ишеевской фауны нижнетатарского подъяруса в Татарстане. Известен по фрагментам черепов, описан Б. П.... -
 Perplexisaurus
Перплексизавры (Perplexisaurus) — род семейства перплексизавровых (Perplexisauridae), происходящий из средней ? — поздней перми Европы. Установлены 2 вида.
Perplexisaurus
Перплексизавры (Perplexisaurus) — род семейства перплексизавровых (Perplexisauridae), происходящий из средней ? — поздней перми Европы. Установлены 2 вида. -
 Moschorhinus
Мосхорин (лат. Moschorhinus) — тероцефал, живший на границе перми и триаса. Принадлежит к семейству Akidnognathidae, близкому к вайтсиидам.
Moschorhinus
Мосхорин (лат. Moschorhinus) — тероцефал, живший на границе перми и триаса. Принадлежит к семейству Akidnognathidae, близкому к вайтсиидам.
-
 penetrat0r
penetrat0r
- 01.06.2018 19:57
-
custodian
- ТС
- 01.06.2018 20:48







