Myosaurus
-
 custodian
custodian
- Вымершие животные
- 13.10.2017 23:10

Миозавр (лат. Myosaurus) — вымерший род дицинодонтов, обнаруженный прежде всего в Антарктиде и Южной Африке.
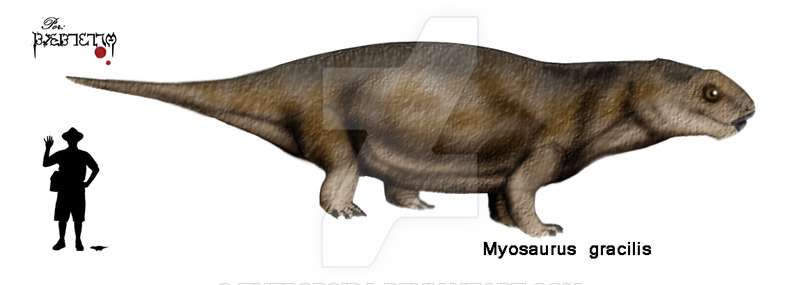
Семейство Myosauridae известно только из образцов окаменелых черепов, семейство отличается от других семейств инфраотряда Dicynodontia большей шириной образцов черепа в интертемпоральной области. Миозавр раньше был членом семейства Endothiodontidae, но после дальнейшего исследования был помещен в свое собственное семейство.

Род Myosaurus известен из раннетриасовой зоны Lystrosaurus Южной Африки и Антарктики по нескольким черепам и ассоциированным фрагментам посткраниального скелета. Несмотря на то что детальное описание остатков Myosaurus было сделано в 1974 г, дополнительное изучение оригинального материала выявило наличие инфраорбитальных отверстий и сопряженной сети кровеносных сосудов на верхнечелюстной кости, которые свидетельствуют о локальном усилении кровоснабжения и иннервации ростральных районов морды и развитии сенсорных зон.
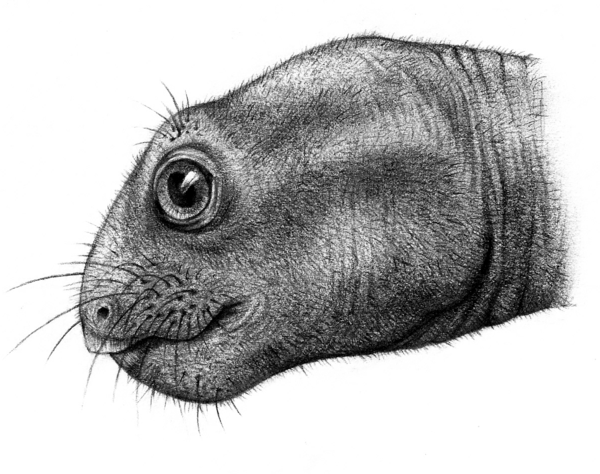
Формирование зон с повышенным кровоснабжением и иннервацией обычно трактовалось как свидетельство наличия вибрисс или сходных образований, выполняющих ту же функцию, что и у современных млекопитающих. Развитие вибрисс также может свидетельствовать и о существовании волосяного покрова – одного из ключевых признаков теплокровности.
Myosaurus gracilis – маленький дицинодонт с черепом длиной около 3,5 см. Морда слабая, ноздри расположены терминально. Орбиты очень большие, ориентированы антеро — латерально. Вентральная поверхность предчелюстной кости вогнута, отсутствуют парные гребни, осложняющие поверхность в передней части, вдоль срединного гребня тянутся два глубоких желобка. Клыковой отросток на верхнечелюстной кости маленький. Лабиальное отверстие отсутствует. Небные кости удлиненные, расширены кпереди , вентральная поверхность костей гладкая. Небное отверстие пронизывает небную кость в передней части. Боковое небное отверстие расположено на внешней стороне небной кости на уровне передних окончаний крыловидных костей. Хоаны разделены длинной и узкой сошниковой костью. Межптеригоидная впадина узкая , ее длина составляет 43% от длины хоан . Наружные крыловидные кости расширены кпереди, изолируя крыловидные кости от контакта с верхнечелюстными костями.
Аналогичное цинодонтам формирование инфраорбитальных отверстий и обширной сети кровеносных сосудов на морде Myosaurus позволяет предполагать развитие сенсорных осязательных зон, которые по аналогии с современными млекопитающими могли быть покрыты вибриссами или вибриссоподобными образованиями. Такое предположение очень хорошо соответствует гипотезе М . Клювера о насекомоядности Myosaurus в отличии от остальных растительноядных дицинодонтов. В пользу этой гипотезы говорят и необычно широкие глазницы, которые на 30% больше сходного по размерам примитивного дицинодонта Emydops. Все эти факты позволяют отнести миозавра к мелким насекомоядным терапсидам, которые вели сумеречный образ жизни. Необычный для дицинодонтов образ жизни этого животного вполне укладывается в рамки послекризисной экосистемы, существовавшей на территории Южной Африки в начале раннего триаса . Во время позднепермского кризиса были разрушены устойчивые прибрежные экосистемы речных пойм, а речная динамика и переработка берегов резко усилились, что в конечном счете привело к формированию узких и неустойчивых прибрежных экотонов с крайне обедненной фауной терапсид . В процессе этих перестроек, вероятно, и произошла диетарная переориентация и специализация мелких дицинодонтов, смогших на короткое время занять нишу мелких насекомоядных процинозухид, из которой они до конца раннего триаса были вытеснены более продвинутыми терапсидами – мелкими тринаксодонами . Последние , помимо вибрисс, имели турбиналии , что свидетельствует о теплокровности и, соответственно, большей активности в холодное время суток. Появление осязательных зон на морде миозавра свидетельствует о том, что один из ключевых признаков млекопитающих – наличие вибрисс, появился конвергентно и почти одновременно во всех группах терапсид в позднепермское – раннетриасовое время. Однако появление и закрепление этого признака в филогенетических линиях происходило не всегда, поэтому его наличие не говорит о настоящей маммальности которая сформировалась значительно позже в результате комбинации дополнительных «отработанных» ранее морфотипов.
Источники:
https://kana-hebi.deviantart.com/art/Myosaurus-gracilis-708099286
https://theropsida.deviantart.com/art/Myosaurus-gracilis-147322473
https://mojcaj.deviantart.com/art/Myosaurus-50761339
https://biarmosuchus.deviantart.com/art/Myosaurus-gracilis-351634096
http://www.sgu.ru/sites/default/files/journals/izvestiya/pdf/2013/12/13/15_2.pdf
http://dinopedia.wikia.com/wiki/Myosaurus
http://www.prehistoric-wildlife.com/species/m/myosaurus.html
Похожие статьи
-
 Repelinosaurus
Репелинозавры (Repelinosaurus) — монотипический род из клады каннемейериеобразных (Kannemeyeriiformes), происходящий из раннего триаса Азии. Единственный установленный вид —...
Repelinosaurus
Репелинозавры (Repelinosaurus) — монотипический род из клады каннемейериеобразных (Kannemeyeriiformes), происходящий из раннего триаса Азии. Единственный установленный вид —... -
 Lisowicia
Lisowicia — это род дицинодонтововых синапсид, которые жили в Польше в позднем норийском или в раннем рэтском ярусах позднего триаса. Будучи размером со слона, это самый...
Lisowicia
Lisowicia — это род дицинодонтововых синапсид, которые жили в Польше в позднем норийском или в раннем рэтском ярусах позднего триаса. Будучи размером со слона, это самый... -
 Stahleckeria
Шталекерия (лат. Stahleckeria) — гигантский дицинодонт середины триаса. Шталекерия относится к группе дицинодонтов-каннемейерид.
Stahleckeria
Шталекерия (лат. Stahleckeria) — гигантский дицинодонт середины триаса. Шталекерия относится к группе дицинодонтов-каннемейерид. -
 Angonisaurus
Angonisaurus — вымерший род каннемериеформного дицинодонта из позднего триаса Африки где-то между 251 млн. лет назад и 199 миллионов лет назад. К этому роду был отнесен только ...
Angonisaurus
Angonisaurus — вымерший род каннемериеформного дицинодонта из позднего триаса Африки где-то между 251 млн. лет назад и 199 миллионов лет назад. К этому роду был отнесен только ...







