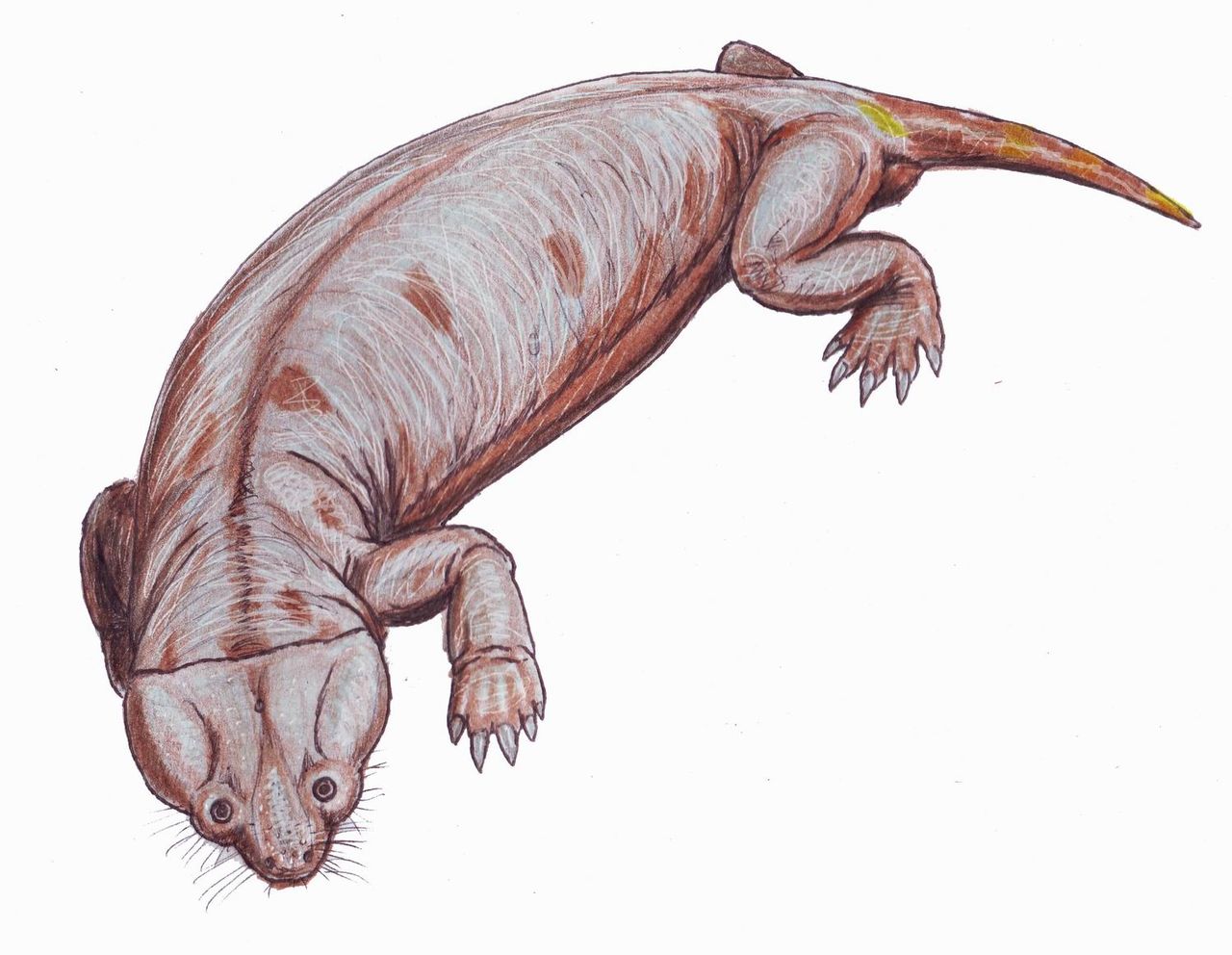
Australobarbarus
-
 custodian
custodian
- Вымершие животные
- 21.02.2019 19:28
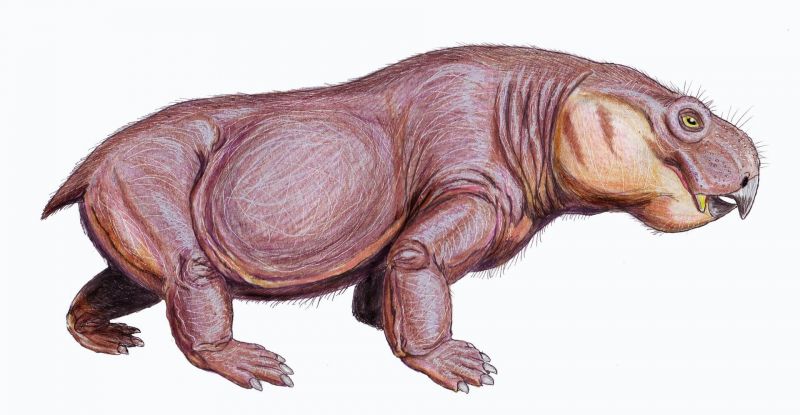
Австралобарбар (лат. Australobarbarus) — род примитивных дицинодонтов из поздней перми Кировской области (котельничская фауна). Род описан А. А. Куркиным в 2000 году, название переводится как «южный варвар». Дело в том, что австралобарбар принадлежит к южноафриканскому семейству (или подсемейству) Cryptodontidae, являясь одним из немногих его представителей в северных фаунах. Череп уплощённый, длиной до 30 см. Скуловые дуги широкие. Клыки-бивни небольшие, могут отсутствовать. Иногда есть остатки челюстных зубов: нёбо могло быть не ороговевшим. По-видимому, питались водной растительностью.
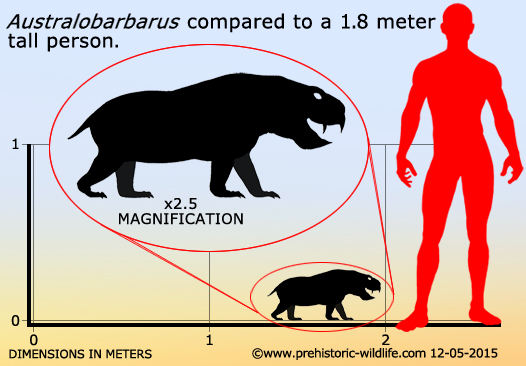
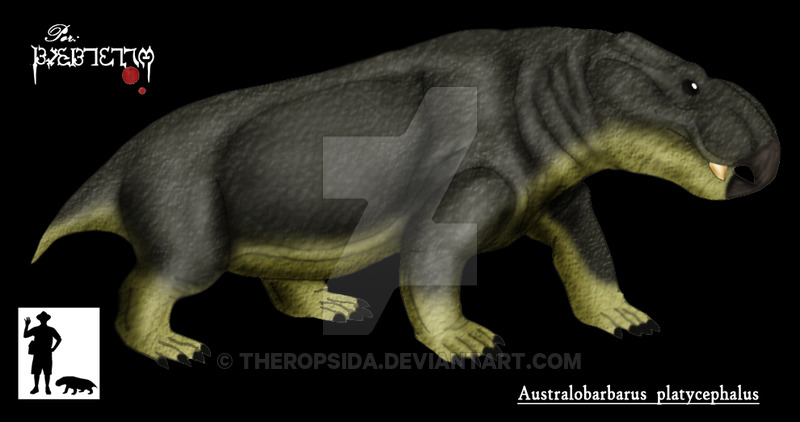
Довольно многочисленны в местонахождении Порт Котельнич (верхнетатарский подъярус, северодвинский горизонт), составляя до 15 % находок. Два вида — A. kotelnitshi (типовой вид) и A. platycephalus. В 2006 году из отложений более позднего ильинского субкомплекса в Татарстане (Семин Овраг) был описан близкий дицинодонт Idelisaurus tatarica.
Фауна котельничского местонахождения (по доминантному таксону Deltavjiatia vjatkensis) наиболее полная из всех позднепермских фаун, представлена преимущественно сухопутным блоком. Водный блок Котельничской фауны полностью отсутствует. Одновременных местонахождений с водным блоком фауны пока не идентифицировано, хотя наверняка были представлены сеймуриаморфами и хрониозухидами. Доминантный блок котельничской фауны был представлен примитивным парейазавром семейства Bradysauridae Deltavjiatia vjatkens, дицинодонтом Australobarbarus platycephalus et kotelnichi, горгонопсом фтинозухом Vjatkogorgon ivachnenkovi. Парейазавры дельтавьятии видимо вели амфибийный образ жизни, не крупные до 1,5 м, могли свободно передвигаться по суши, возможно, были способны к далеким миграциям. Об этом свидетельствует, обнаружение комплекса следовых дорожек, предположительно принадлежащих парейазаврам близким по размерам с Deltavjiatia vjatkensis. (местонахождение Устье Стрельны, р. Сухона Вологодская область). По форме зубного аппарата можно предположить о питании парейазавров мягкой пищей, водными матами состоящими из растительной биомассы водорослей, простейших и ракообразных остракод. О предположительном питании парейазавров ракообразными остракодами писал Д.Л. Сумин в 2009 году. В литературе не встречается упоминаний о стирании зубов парейазавров. Но в коллекции ВПМ имеется зуб парейазавра Proelginia cf. Permiana (местонахождения Чижевская линза) верхний зубчик которого прижизненно стерт противопоставленным зубом. Поликупсидный зуб парейазавра (у дельтавьятий 11 зубчиков видимо приспособлены не только для отцеживания водной биомассы, но и для срезания мягких листьев водорослей. Таким образом можно интерпретировать образ жизни парейазавра как крупную наземную рептилию, с питанием в мелких озерных водоемах, водными водорослями и планктоном, но передвигавшуюся по суши довольно активно. Данная реконструкция образа жизни подходит только для наиболее примитивных Восточно-Европейских парейазавров Deltavjiatia vjatkensi. Для более продвинутых, более поздних парейазавров Proelginia permiana, Scutosaurus karpinski, S. Itilensis, S. Tuberculatus М.Ф. Ивахненко предполагает облигатно водный образ жизни, без возможности передвигаться по суше, видимо с узкой специализацией к такому образу жизни связано вымирание скутозавров незадолго до конца перми. В поздней терминальной перми считается, происходило увеличении аридизации, осушение ландшафтов, исчезновение мелких мореподобных озер, соответственно исчезают обитатели этих озер скутозавры и специализированные на их охоте крупные горгонопсы Иностранцевии.

Другим крупным растительноядным элементом котельничской фауны являются дицинодонты. Из местонахождения Порт Котельнич описано два вида одного рода Australobarbarus platycephalus et kotelnichi относимых к экологической группе наземных крупных растительноядных животных. Котельничские дицнодноты рептилии среднего размерного класса длиной до метра, с относительно крупной головой. О пищевых пристрастиях дицинодонтов до сих пор ведутся споры и предполагается питание плодами, грибами, беспозвоночными и даже падалью. Также подвержен спорам вопрос о функциональной нагрузке клыков. Последние работы по южноафриканским дицинодонтам опровергают использование их для рытья почвы. Последние работы (Куркин А.А., 2012) предполагают использование клыков как мощных тисков или зацепа для вытягивания сочных корневищ из земли. Это косвенно можно потвердить наличием недалеко от местонахождения Порт Котельнич палеопочвенных профилей с наличием корневых отпечатков типа Rhadycites sp.в виде глубокоуходящего вглубь единого корня. И их отсутствием в типовом местонахождении парейазавров возле д. Боровики, в почвенных профилях присутствуют корневые системы другого типа в виде разветвленной сети. Таким образом мы относим котельничских дицинодонтов к группе наземных фитофагов.
Вершиной пищевой цепи в котельничской фауне был горгонопс фтинозухид Vjatkogorgon ivachnenkovi средних размеров хищник, но с довольно грацильным скелетом, позволявшим ему довольно активно передвигаться на суше, в отличие от парейазавров, у него произошла редукция задних туловищных ребер и формирование поясничного отдела. Как у всех горгонопсов, у Вяткогоргона довольно хорошо развиты клыки, и сложный нижнечелюстной сустав с дополнительным кинетизмом черепа, что позволяло открывать пасть на довольно большой угол. Данный тип челюстного соединения наиболее полно реализовался у поздних, специализированных, горгонопсов типа иностранцевии. У Вяткозуха можно предполагать еще не сильноразвитую специализацию охоты на парейазавров. Поскольку останки горгонопсов найдены как у д. Боровики так и у. г. Котельнич в одних слоях и с парейазаврами и с дицинодонтами можно предположить, что вяткогоргон питался обоими растительноядными формами. О способе охоты можно предположить как о засадном типе, исключавшем длительную погоню и нападение.
Большинство дицинодонтов, скорее всего, представляет собой в большей или меньшей степени специализированных фитофагов. В поддержку гипотезы о том, что фитофагия являлась генеральной эволюционной линией дицинодонтов, можно привести ряд общебиологических доводов. Прежде всего, это – многочисленность остатков дицинодонтов во многих местонахождениях, что, вероятно, отражает и многочисленность этих животных в пермских биоценозах. Подобная обильность по сравнению с явными хищниками свидетельствует в пользу низкого положения дицинодонтов в трофической пирамиде сообщества и соответствует многочисленности фитофагов в современных экосистемах.
Необходимо различать фитофагию на основе мягких частей растений, например, генеративных частей или плодов, что, как правило, не требует интенсивной предварительной обработки пищи челюстями и может не приводить к формированию специфических морфофункциональных адаптаций, и фитофагию на основе частей растений с высоким содержанием жестких волокон – «high-fibre plant material» (Sues, Reisz, 1998, p. 141). Видимо, именно последний тип питания и формирует характерный облик фитофага со специфическими морфофункциональными адаптациями. Следует различать основные объекты, к питанию которыми животные имеют специфические морфологические приспособления, и возможные факультативные объекты, которые, впрочем, могут составлять значительную часть рациона. Разные авторы предполагали различные объекты питания для дицинодонтов: плоды, грибы, беспозвоночные, падаль (Sushkin, 1926; Cox, 1972; Kemp, 1982; Hotton, 1986) и, таким образом, некоторые из дицинодон- тов рассматривались как консументы второго порядка.
Предположения о возможном специфическом использовании бивней для рытья лишены всяких оснований. Исследуя поверхности стирания бивней у верхнепермских дицинодонтов, С. Хоттон (Hotton, 1986) приходит к выводу, что только у нескольких родов (Dicynodon, Lystrosaurus, Diictodon и, возможно, Kingoria), и только при максимальном развитии бивней, когда при закрытой и полностью отведенной назад нижней челюсти не более трети длины бивня выступает за пределы челюстей, они могли бы использоваться при копании. Вероятно, в первую очередь при использовании рытья для добывании пищи использовались конечности, а не зубы (Hotton, 1986), к чему наибольшие приспособления (исключая цистецефалид) обнаруживает Diictodon. Одним из возможных объяснений функционального значения бивней дицинодонтов может служить предположение об использовании их в качестве специфического орудия захвата и удержания объекта в клещевом замке, образованном, при смыкании челюстей, бивнями (или клыкообразными выростами) и нижней челюстью. Такой механизм мог бы оказаться весьма эффективным при питании, например, слабо и неглубоко укорененными подземными частями (побегами, корневищами) растений, позволяя извлекать из почвы значительные фрагменты подземных частей растений для последующей обработки. В этом случае копание осуществлялось не бивнями, а передней частью головы, покрытой чехлом ороговевавшей кожи, в мягком, скорее всего влажном грунте и на небольшую глубину. Подобное «копание» можно весьма отдаленно сравнить с «копанием» кабанов и диких свиней. Подобные адаптации могли использоваться в условиях субаквальных обстановок (Ивахненко, 2001) с доминированием плауновидных, хвощей и папоротников (Наугольных, 2004).
Позднепермские дицинодонты Восточной Европы, видимо, представляли собой в среднем и крупном размерном классе специализированных «обкусывателей» сравнительно мягкой низкорослой растительности (Australobarbarus, Idelesaurus), менее специализированных по объекту питания «собирателей» (Fortunodon, Delectosaurus), а в мелком размерном классе – неспециализированных фитофагов и, возможно, омнифагов или инвертеброфагов (Elph, Interpresosaurus). Отдельный тип специализации представляет собой Vivaxosaurus, приобретающий черты крупного фитофага (каннемейероидные черты), в том числе и в постановке головы (клинориния черепа).
На рисунке ниже представлены Deltavjatia, Vjatkogorgon, Australobarbarus.

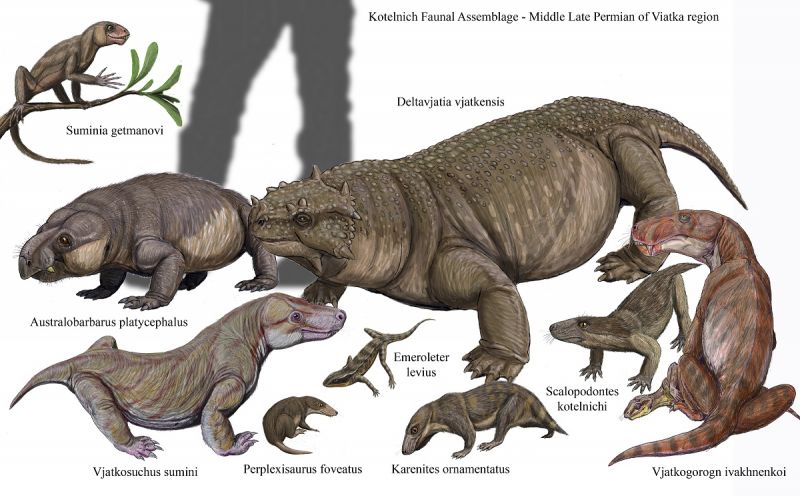
На рисунке ниже представлена фауна Котельнича (Deltavjiatia, Suminia,Australobarbarus, Emeroleter, Vjatkogorgon, Vjatkosuchus, Karenites, Perplexisaurus, Scalopodontes).
Источники:
http://www.prehistoric-wildlife.com/species/a/australobarbarus.html
https://www.ammonit.ru/foto/42290.htm
https://museum-noyabrsk.ru/collections/skelet-dicinodonta-australobarbarus-kotelnichi-slepok
http://www.paleofile.com/Dicynodont/Australobarbarus.asp
https://www.deviantart.com/abelov2014/art/Deltavjatia-Vjatkogorogn-Australobarbarus-526708080
https://uk.wikipedia.org/wiki/Australobarbarus
http://dis.podelise.ru/docs/index-2804.html
https://www.deviantart.com/romanyevseyev/art/Australobarbarus-kotelnitshi-461521178
https://www.deviantart.com/theropsida/art/Australobarbarus-platycephalu-146588615
https://en.wikipedia.org/wiki/Australobarbarus

Похожие статьи
-
 Oudenodon
Oudenodon — род вымерших дицинодонтов из конца перми Южной Африки. Обнаружен в Замбии, Карру и на Мадагаскаре.
Oudenodon
Oudenodon — род вымерших дицинодонтов из конца перми Южной Африки. Обнаружен в Замбии, Карру и на Мадагаскаре. -
 Emydops
Эмидопс (лат. Emydops) — род небольших дицинодонтов. Открыт в 1912 году. Обитал с середины по конец пермского периода в Южной Африке. Название эмидопсу в 1912 году дал...
Emydops
Эмидопс (лат. Emydops) — род небольших дицинодонтов. Открыт в 1912 году. Обитал с середины по конец пермского периода в Южной Африке. Название эмидопсу в 1912 году дал... -
 Endothiodon
Эндотиодон (лат. Endothiodon) — дицинодонт середины позднепермской эпохи.
Endothiodon
Эндотиодон (лат. Endothiodon) — дицинодонт середины позднепермской эпохи. -
 Cistecephalus
Кистецефал (лат. Cistecephalus, от др.-греч. κύστις «коробка» и κεφᾰλή «голова») — мелкий дицинодонт позднепермской эпохи. Роющее животное, своего рода крот пермского периода. ...
Cistecephalus
Кистецефал (лат. Cistecephalus, от др.-греч. κύστις «коробка» и κεφᾰλή «голова») — мелкий дицинодонт позднепермской эпохи. Роющее животное, своего рода крот пермского периода. ...







